Cell Communication
Introduction
Cell communication allows one cell to change the behavior of another cell. Cells communicate using a language that is detected by cells and generates a response. The response usually involves a change in one or more biochemical pathways that alter the behavior of a cell. Cells use the same mechanisms to detect changes in their external environment and respond appropriately.

Cell Communication Language
The language of cells consists of a variety of different molecules that differ in size and chemical composition. Some are hydrophilic: amino acids and their derivatives, proteins and peptides. These molecules must bind a receptor in the cell membrane. Other molecules are hydrophobic: gases and steroids. These can readily diffuse across the cell membrane and interact with intracellular receptors.

Interpretation of Cellular Language
Much like our words, the molecules that mediate cell communication can be interpreted differently by different types of cells. The same molecule can stimulate different responses depending on the type of cell. For example, acetylcholine is a neurotransmitter that stimulates contraction in skeletal muscle, but in cardiac muscle cells acetylcholine decreases the rate of contraction. The difference between skeletal and cardiac muscle cells is in how they are wired biochemically to respond to acetylcholine.
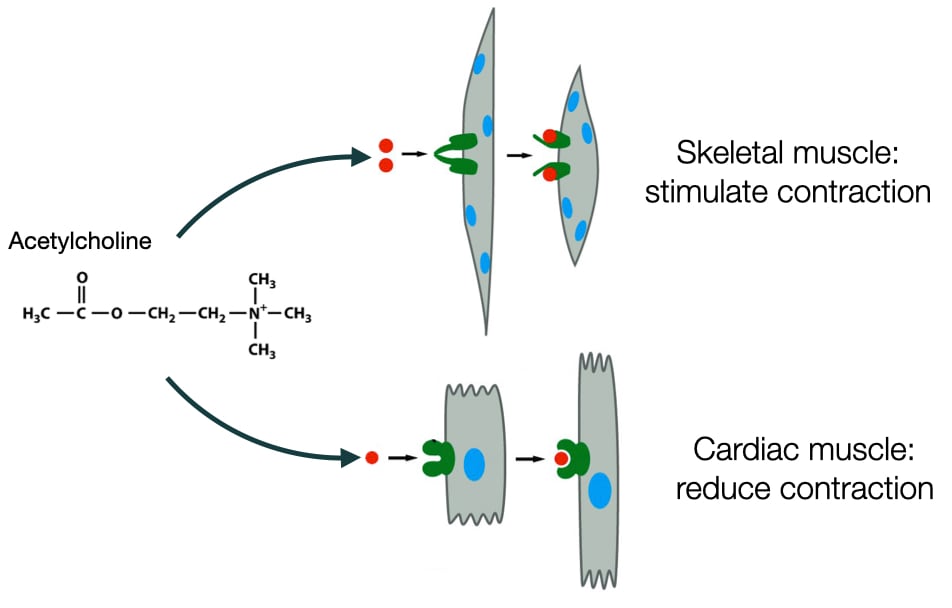
The ability of cells in different tissues and organs to respond differently to same signaling molecules, means that one molecule can generate an integrated response in the body that involves multiple organs. For example, epinephrine affects cells in a variety of organs and tissues to produce a flight or fight response in an organism. Cells respond differently to epinephrine depending on which organ or tissue they reside. The biochemical wiring in these cells differs to produce to different responses to epinephrine.

Fast and Slow Responses in Cell Communication
A cell’s response to a signaling molecule can take different lengths of times. A fast response is achieved by altering the activities of existing proteins that lead directly to changes on existing biochemical pathways. The response is fast because the signaling event does not require the synthesis of new protein but instead modifies existing proteins and other macromolecules to alter their conformation and behavior. Fast responses are usually reversible as once the signaling molecule is removed the modifications induced by the signaling pathway can be quickly returned to normal.
Cells also respond to signaling molecules by altering gene expression to increase or decrease the concentrations of specific proteins. Changing the concentration of a protein can alter the rate of biochemical reactions that lead to changes in cell behavior. These pathways are slow because it requires time to build up or remove protein to cause a change in cell behavior. The slow response often generates long-term changes in cells such as differentiation into a specific type of cell or cell division and may not be reversible.
Some signaling molecules elicit both types of pathways in cells.

Type of Cell Communication
Communication between cells can be classified based on the distance over which the communication takes place.
Paracrine signaling
In paracrine signaling, the signal molecule released by one cell only affects cells in the surrounding area. Importantly, the signaling molecule does not enter blood stream. The extracellular matrix surrounds the cells and restricts the diffusion of the signaling molecule. A special type of paracrine signaling is autocrine in which the cell that produces the signaling molecule also responds to that signaling molecule. Paracrine signaling is important during development as it allows organisms to produce cells that will develop different functions. A signaling molecule (morphogen) released from one point will diffuse away from that point creating a gradient of morphogen concentration. Cells will respond differently depending on the concentration of the morphogen.

Endocrine Signaling
Endocrine signaling involves cells located in different parts of an organism. Signaling molecules secreted by one cell enter blood stream and travel through blood stream to other parts of body. Cells throughout the body have an opportunity to detect and respond to the signaling molecule. Signaling molecules in endocrine signaling are called hormones.

Signaling Through Direct Contact
A third type of signaling involves direct interaction between cells. Proteins in the cell membranes of different cells bind each other and the interaction between these proteins alters cell behavior. For example, a T-cell uses its receptor to recognize antigen on the surface of antigen presenting cell. Another form of this type of signaling is the communication between neurons and their target cells. Neurons make contact with their target cell and but don’t communicate via these contacts. Instead, they release neurotransmitter at these contact points to affect their target cell.

Strength of Interaction Between Receptor and Signaling Molecule
The different types of signaling differ in the strength of interaction between the signaling molecule and the receptor for that molecule. In endocrine signaling, the hormones are present at low concentration because they are distributed throughout the body. In addition, cells are often bathed in a variety of different hormones and other signaling molecules. Thus, the receptors on cells have to distinguish between several different molecules at low concentration. This requires a high affinity interaction between the receptor and its hormone.
In paracrine signaling or signaling via neurons, the signaling molecule is usually present at higher concentration and is sometimes the only signaling molecule present (neurotransmission). Consequently, the strength of the interaction between the receptor and its signaling molecule is usually lower than in endocrine signaling.
The importance of the strength of interaction between receptor and its signaling molecule comes when the cell wants to turn off its response to the signaling molecule. A low affinity interaction between a signaling molecule and receptor means the signaling molecule more easily dissociates from the receptor. By releasing the signaling molecule more readily, signaling pathways are inactivated quicker and the response to the signaling molecule ends sooner. A high affinity interaction prolongs the association between a signaling molecule and receptor. Cells cannot rely on the quick dissociation of signaling molecule and receptor to turn off signaling pathways. Instead, cells often degrade the receptor to reduce its concentration and inactive signalin

Mechanisms of Detecting and Responding to Signaling Molecules
Cell Communication by Steroids
One of the simplest signaling pathways involves steroid hormones and their mechanism of activating expression of specific genes. Steroids usually function as hormones. They are produced by specific cells in one region of the body and use the circulatory system to reach their target cells. All steroids are synthesized from cholesterol and are hydrophobic molecules that can diffuse across the cell membrane. The receptors for steroids are transcription factors. Some of these receptors are kept in the cytosol by a chaperone that hides the nuclear localization sequence (NLS) in the receptor. When the receptor binds steroid, it releases the chaperone, exposing the NLS. Once imported into the nucleus, the receptor can activate transcription of specific genes.

How Steroids Alter Cell Behavior
Steroids alter cell behavior through two rounds of transcription. As mentioned, steroids bind their receptors which are transcription factors. These transcription factors, when bound to steroid, increase the expression of a set of genes called primary response genes. Primary response genes contain a regulatory sequence called a sterol-response element where the steroid receptors bind to activate transcription. The primary response genes encode proteins that function as transcription factors. Some these transcription factor increases the expression of a set of genes called secondary response genes. These genes encode proteins that alter cell behavior. Some of the transcription factors encoded by the primary response genes inhibit the expression of the primary response genes. This is an example of negative feedback. Thus, steroids not only lead to the activation of genes that change cell behavior but also genes that decrease a cell's response to the steroid.

Cell Communication via Ion Channels
Another simple way signaling molecules can alter cell behavior is through depolarization of the cell membrane to trigger an action potential. Some receptors are ion channels. In the absence of ligand (signaling molecule), the channel is closed. When bound to the ligand, the channel opens allowing ion to diffuse across the cell membrane. The acetylcholine receptor is an example of a ligand-gated ion channel which depolarizers the cell membrane of skeletal muscle cells.

Signal Transduction
Many signaling molecules cannot diffuse across the cell membrane nor do they bind ligand-gated ion channels. Instead, these signaling molecules require proteins to transduce their presence from outside the cell to proteins inside the cell. The pathways that transmit this information are called signal transduction pathways.
Cell Membrane Receptors
Receptors are critical to signal transduction pathways. The receptors are usually integral membrane proteins in the cell membrane. The portion of the protein that faces the external environment contains a domain which binds to a specific signaling molecule. Domains in the protein that reside in the cytosol interact with the downstream components of the signaling pathway. When the receptor binds a signaling molecule, it undergoes a conformational change that allows the receptor to activate the signaling pathway and alter cell behavior.

Downstream Effectors of Cell Membrane Receptors
Several components relay the binding state of the receptor to the cell machinery (metabolism, gene expression, morphology). Two of the most common components are GTP-binding proteins and kinases. Both function as switches that transmit the ligand-binding state of the receptor to the other components in the signal transduction pathway.
GTP-binding Proteins
Signaling pathways that are activated by receptors often use biochemical switches to indicate the bound or unbound state of the receptor. GTP-binding proteins are example of a biochemical switch and are used in many signaling pathways. GTP-binding proteins are inactive when bound to GDP and active when bound to GTP. To cycle between these states, GTP-binding proteins bind GTP and then slowly hydrolyze the GTP to GDP and phosphate. Phosphate is released to generate a GTP-binding protein with GDP. GTP-binding proteins release GDP slowly and because the concentration of GTP is higher than GDP in the cytosol, GTP-binding proteins are more likely to bind GTP and restart the cycle.

Because the GTP hydrolysis and GDP-release rates are slow, signaling pathways increase the rates of the steps to switch the GTP-binding protein between the GTP-bound (active) state and GDP-bound (inactive state). Two proteins convert GTP-binding proteins between the GTP-bound and GDP-bound states. Guanine nucleotide exchange factors (GEFs) catalyze the release of GDP allowing the protein to bind GTP. Thus, GEFs turn on GTP-binding proteins. In contrast, GTPase-activating proteins (GAPs) increases the rate of GTP hydrolysis in GTP-binding proteins converting them to the GDP-bound or off state.
Kinases
Another type of switch in signaling pathways involves phosphorylation. Kinases are proteins that add phosphates to proteins. The presence of phosphate will alter the activity, stability and/or location of the protein. Phosphatases remove phosphate groups. Usually receptors and signaling pathways activate kinases to phosphorylate specific proteins which either continues the signaling pathway or alters the behavior of the cell. For example, phosphorylation of an enzyme could increase its activity to stimulate a biochemical pathway or phosphorylation could mark a protein for degradation to turn off a biochemical pathway.

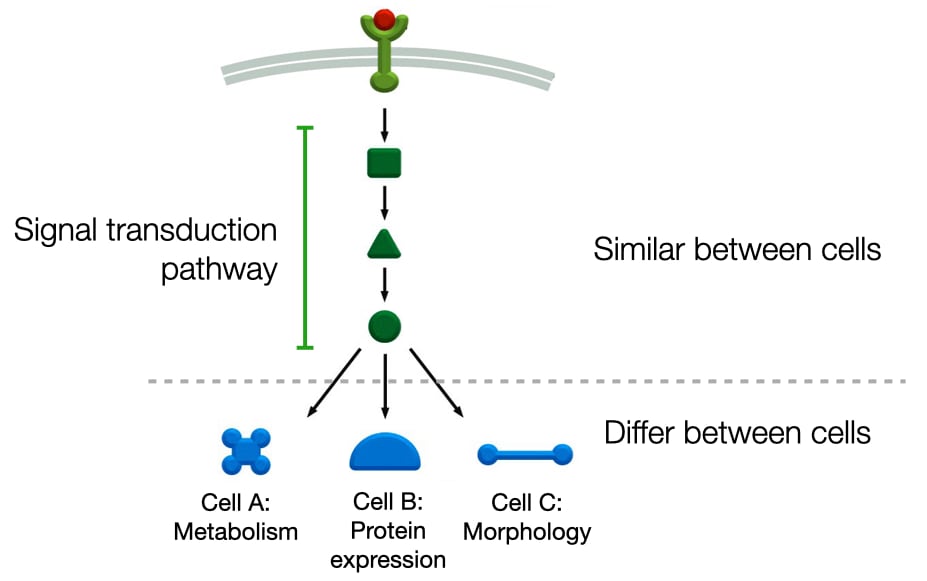
Signal Transduction Pathways
In addition, to GTP-binding proteins and kinases, signaling pathways can include several other types of proteins to relay the binding state of the receptor to the cell machinery (metabolism, gene expression, morphology). These same components are often used in different cells and connected to different receptors. For a cell to generate a different response to a signaling molecule, the components of the signaling pathway are connected to a specific biochemical pathway (e.g. metabolic pathway, gene expression pathway or cell structure pathway). As a result, different cells can produce different responses to the same signaling molecule.
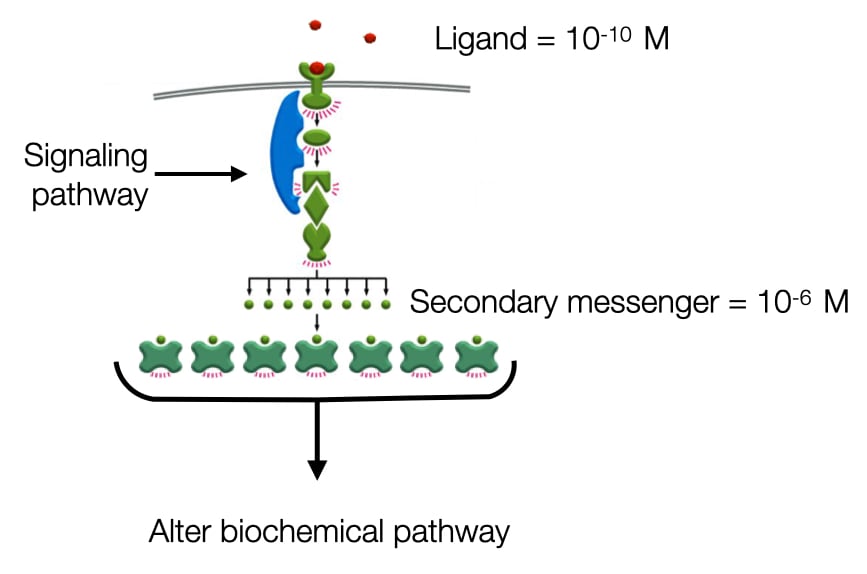
Amplification of Signals
One benefit of having many components between signaling molecule and cell machinery is it allows for amplification of signal. Many signaling molecules are present at low concentration (e.g. 10<sup>-10</sup> M). Cells contain only about 1000 receptors for a given ligand but they need to activate hundreds of thousands or millions of proteins to alter cell behavior. To amplify the signal signaling pathways activate enzymes each of which is capable of producing many copies of an active molecule or protein. For example, adenlylyl cyclase is an enzyme that produces cAMP and can increase the cytosolic concentration of cAMP to 10<sup>-6</sup> M which is a 10000-fold higher concentration than the signaling molecule outside the cell. cAMP can activate specific proteins in the cytosol to continue the signaling pathway. cAMP is an example of the secondary messenger. The signaling molecule outside the cell is the primary messenger..
Inactivating Receptors
In addition to responding to ligands, cells also regulate the strength and duration of their response to a ligand. If a cell's response to a ligand is too strong or remains active for too long, the cell can become damaged. In some instances, this can lead to cell death. Therefore, signaling pathways will usually trigger reactions that reduce the strength of a cell's response to a ligand (negative feedback). These reactions will often reduce the number of receptors in the plasma membrane or prevent the receptor from activating a cellular response.

Positive and Negative Feedback
Signaling pathways usually employ two types of feedback to regulate the strength of the pathway. Most commonly, signaling pathways use negative feedback to limit the strength of the pathway. An example of negative feedback is when a receptor turns on an enzyme by phosphorylating the enzyme. The enzyme activates other components in the signaling pathway and also a protein that inactivates the enzyme. In this example the inactivating protein would be a phosphatase that removes the phosphate from the enzyme to inactive the enzyme.
The graphs compare the strengths of a response generated by pathways without and with negative feedback. Without negative feedback, the strength of the response remains high while the stimulating signal is present. With negative feedback, the response has an initial strong output which is then reduced by negative feedback to a lower level. Negative feedback is also capable of generating oscillations in the strength of the response which can generate cell behaviors that change over time.

Positive feedback is less commonly used in signaling pathways because the response to the signal becomes independent of the presence of the signal. An example of positive feedback involves a signal that activates an enzyme through phosphorylation of the enzyme. Once activated, the enzyme not only turns on the downstream pathway but also activates more of itself through autophosphorylation (the enzyme can phosphorylate itself). Note that once the enzyme has been activated, it can be generate more active enzyme by phosphorylating other copies of the enzyme. Even if the initial signal is removed, the presence of phosphorylated enzyme ensures that the enzyme remains active.

Type of Signaling Pathways
Although there are many different signal transduction pathways, there are a few very common pathways. One involves a family of proteins called G-protein coupled receptors (GPCR) that recognize a wide variety of molecules and are linked intracellularly to a class of GTP-binding proteins called a heterotrimeric GTP-binding protein. The other common signal transduction pathway uses tyrosine kinase receptors.
Heterotrimeric GTP-binding Proteins
Heterotrimeric G-proteins are a large family of GTP-binding proteins that consist of three separate proteins. The alpha subunit binds and hydrolyzes GTP and the beta-gamma subunits serve as accessory proteins that keep the alpha subunit bound to GDP. The beta-gamma subunits can also have roles in signaling pathways separate from the alpha subunit. The heterotrimeric G-proteins are kept at the cell membrane via short hydrocarbon chains that are covalently linked to the alpha and gamma subunits and insert into the lipid bilayer.
Signaling through heterotrimeric G proteins usually occurs via receptors that have seven transmembrane domains and are referred to as G-protein coupled receptors. The class comprise a large family of receptor proteins that are found in cells of most tissues and organs. Upon binding ligand, the receptor undergoes a conformational change that activates its guanine nucleotide exchange domain. The domain catalyzes nucleotide exchange on an associated alpha subunit, leading to an alpha subunit bound to GTP. In the GTP-binding state the alpha subunit dissociates from the beta-gamma subunits. Both are capable of regulating downstream signaling pathways.

Adenylyl Cyclase
One of the major downstream effectors of G alpha (Gα) is adenylyl cyclase. Adenylyl cyclase is cell-membrane bound protein that catalyzes the conversion of ATP to cyclic AMP. It increases the cytoplasmic concentration of cAMP 20 fold in a few seconds. cAMP is secondary message that will trigger further downstream signaling events. Two different types of alpha subunits regulate the activity of adenylyl cyclase. Alpha stimulatory (Gαs) subunits increase the activity of adenylyl cyclase to produce more cAMP. Alpha inhibitory subunits (Gαi) inhibit adenylyl cyclase to turn down production of cAMP.

Protein Kinase A
Most of the effects of increased cAMP are mediated by protein kinase A. Protein kinase A is a tetramer of two catalytic subunits and two regulatory subunits. In the tetramer form, the regulatory subunits inhibit the activity of catalytic subunits. cAMP binds the regulatory subunits and causes them to dissociate from the catalytic subunits. Once freed, the catalytic subunits are active and can phosphorylate target proteins. Protein kinase A has many downstream targets. For example in some cells, it increases the cytosolic concentration of glucose by activating glycogen phosphorylase to break down glycogen while inactivating glycogen synthase. In other cells protein kinase A phosphorylates the transcription factor CREB that binds to DNA and activates transcription of specific genes.

Phosphodiesterase
Cells must turn off signals to prevent over activation. To limit the activity of enzymes turned on by cAMP, cells express a consistent level of phosphodiesterase that converts cAMP into 5’-AMP. In unstimulated cells, phosphodiesterase keeps cytosolic concentrations of cAMP at a low level so there are few active protein kinase A proteins. Activation of an appropriate G-protein coupled receptor increases activity of adenylyl cyclase. The increase in active adenylyl cyclase produces more cAMP than phosphodiesterase can process, increasing the concentration of cAMP. When adenylyl cyclase is turned off, phosphodiesterase returns cAMP concentrations to resting levels.

In addition to converting ATP to cAMP, adenylyl cyclase functions as a GTPase-activating protein for G alpha subunits. For example, when a Gα which is in a GTP-bound state interacts with adenylyl cyclase, the interaction not only activates adenylyl cyclase but it also accelerates the rate of GDP hydrolysis by Gα. This mechanism both activates downstream pathways and provides negative feedback to limit the strength of the pathway.
Signaling via Phospholipids
Another pathway activated by Gα subunits involves a special set of phospholipids called phosphatidylinositols. Similar to other phospholipids, phosphatidylinositols have fatty acids that span the inner leaflet of the cell membrane but also contain an inositol head group that can be modified by phosphate at different positions. The different combinations of phosphates on inositol are recognized by different proteins and lead to unique cell responses. Kinases can generate the different phosphatidylinositols in a cell membrane.

Phospholipase C
Some G alpha subunits activate phospholipase C, an enzyme in the cell membrane. Phospholipase C cleaves the head groups from certain phospholipids. For example, phospholipase C cleaves the head group phosphatidylinositol 4,5-phosphate to produce diacylglycerol and inositol-1,4,5 triphosphate (IP3).

IP3 and Protein Kinase C
IP3 and diacyl glycerol work together to activate protein kinase C. IP3 diffuses to the endoplasmic reticulum where it binds a calcium-channels called the IP3 receptor. Binding to IP3 opens the channel to release calcium into the cytoplasm. Protein kinase C in the cytosol binds calcium and traffics to cell membrane where it binds diacylglycerol, leading to its activation. Protein kinase C has many cellular targets, and there are different classes of protein kinase C with different targets.

Receptor Tyrosine Kinases
Receptor tyrosine kinases are cell membrane proteins and are inactive in the absence of their ligand because they exist as monomers that don’t interact with targets efficiently. When bound to ligand, the receptors dimerize. Dimerization allows the kinase domain in one receptor to phosphorylate its partner receptor and vice versa in a process called cross-phosphorylation.

Recruitment of Proteins
The phosphorylated cytosolic tails of receptor tyrosine kinases are recognized by different downstream signaling proteins These proteins have domains that recognize portion of the receptor and a phosphate. The phosphorylated receptor often brings together proteins that function in a common signaling pathway. As a result, the enzymes in the pathway are closer to their substrates which accelerates the reaction in the pathway. In the absence of phosphorylated receptor, enzymes and substrates are at low concentration in cytosol and the reactions are very inefficient.

Recruitment to the Cell Membrane
An example of a receptor tyrosine kinase pathway involves recruiting proteins to the cell membrane. When cross-phosphorylated receptor tyrosine kinases recruit proteins that alter the composition of the cell membrane. In the example shown, the phosphorylated receptor bind PI3 kinase. Binding of PI3 kinase leads to its activation and brings it closer to its substrates phosphatidylinositol-4,5-phosphate (PIP2) in the cell membrane. PI3 kinase phosphorylates PIP2 to convert it to phosphatidylinositol-3,4,5-phosphate (PIP3). A class of proteins that contain a PH domain binds PIP3 in the cell membrane. In the pathway below, two different kinases with PH domains are recruited to the cell membrane by PIP3. One kinase phosphorylates the other activating the second kinase. The second kinase can now activate its downstream targets. Without PIP3 in the cell membrane, the two kinases would exist in the cytosol at low concentrations and the probability of interacting with each other is low so the activation of the second kinase is very slow. PIP3 accelerates this pathway by concentrating both kinases at the cell membrane.

Cells can turn off this pathway by reducing the concentration of PIP3 in the cell membrane. For example, a protein called SHIP removes a phosphate from PIP3 to produce PIP2.
MAP Kinase Pathways
Another pathway that receptor tyrosine kinases work through involves a series of cytosolic kinases called MAP kinases. MAP stands for mitogen activated protein because the kinases are often activated by a class of signaling molecules called mitogens which stimulated cell division. In this pathway, the phosphorylated tyrosine kinase recruits two proteins, one of which functions as a guanine nucleotide exchange factor. The guanine nucleotide exchange factor catalyzes a small GTP-binding protein, called Ras, to release GDP and bind GTP. When bound to GTP, Ras changes conformation and activates MAP kinase kinase kinase. MAP kinase kinase kinase then phosphorylates a protein called MAP kinase kinase to activate it. Then MAP kinase kinase phosphorylates MAP kinase to activate it. The active MAP kinase can then phosphorylate enzymes in biochemical pathways that alter cell behavior.

Why does the pathway employ a series of kinases? One explanation is that it can amplify the signal. Because a kinase at each level can phosphorylate several kinases at the next level, the concentration of active MAP kinases is much higher than the concentration of MAP kinase kinase kinase. Second, the multiple kinase provide entry points for regulatory proteins to control the strength of the response.
Inactivating Receptors
If signal transduction pathways are active for too long, cells become damaged and eventually undergo apoptosis. One mechanism to decrease the strength of a signal transduction pathway is to prevent the receptor from activating the downstream components in its pathway.
Receptors can be inactivated by masking the portion of the receptor that activates downstream components of a signaling pathway. Often signal transduction pathways will not only alter specific biochemical pathways but also activate proteins that decrease the strength of the pathway. In the image below, the signal transduction pathway activates a kinase that phosphorylates the receptor which had turned on the signal transduction pathway.

For receptor that bind tightly to their ligand, cells often have to degrade the receptor to turn off the signaling reactions. Receptors are endocytosed into clathrin coated pits. After clathrin is removed, membrane surrounding vesicle undergoes invagination to form internal vesicles that contain receptors. The multi vesicular body will fuse with the lysosome where the receptor is degraded.

Integrated Signaling Pathways
Cells are bathed in variety of different signaling molecules and the behavior of a cell is usually depends upon how a cell detects and responds to these signaling molecules. Altering cell behavior often requires changing the amounts of more than one type of signaling molecule.

The different signaling molecules that surround cells interact with different receptors, and these receptors will stimulate different signal transduction pathways. Often, these pathways will interact with each other by sharing components or when a component in one pathway affects the activity of a component in another pathway. Different signaling molecules can work synergistically to increase the strength of a signal or competitively to reduce the strength of a signal. Thus, a combination of signaling molecules can generate an integrated and tailored response in cells that depends upon which signaling pathways are activated.
