Muscle
Introduction
Muscles are multicellular contractile units. They are divided into three types:
- Skeletal muscle
- Cardiac muscle
- Smooth muscle
As you read about each type of muscle, think about the similarities and differences between them in terms of structure and function.
Skeletal Muscle
Skeletal muscle is mainly responsible for the movement of the skeleton, but is also found in organs such as the globe of the eye and the tongue. It is a voluntary muscle, and therefore under conscious control. Skeletal muscle is specialized for rapid and forceful contraction of short duration.
Organization of Skeletal Muscle
Skeletal muscle is made up of elongated cylindrical multinucleate cells, also called muscle fibers, that span the length of the muscle. Each muscle cell is surrounded by connective tissue called endomysium. Endomysium is similar in composition to basement membrane that underlies epithelia. Muscle cells are grouped together into fasciculi and surrounded by a connective tissue called perimysium that is rich in collagen fibers. Finally, several fasciculi are bundled to form the whole muscle. The whole muscle is surrounded by a connective tissue layer called the epimysium. Blood vessels and nerves travel through the epimysium, perimysium and endomysium to reach the individual muscle fibers.
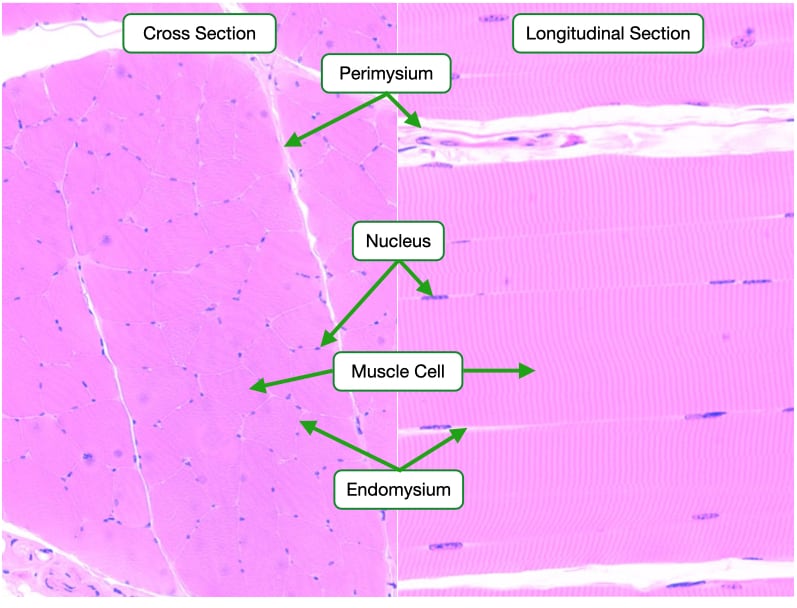
Skeletal muscle cells are easily identifiable in histological samples. In cross section, skeletal muscle cells appear as large cells with their nuclei located to one side. In longitudinal section, the striations identify skeletal muscle cells. The cytoplasm of skeletal muscle cells is organized into long cables called myofibrils that span the length of the cell. Myofibrils are sectioned longitudinally into sarcomeres that give skeletal muscle cells their striated appearance
Sarcomere Electron Micrograph
Sarcomeres are the contractile unit of myofibrils. Sarcomeres are defined by Z-discs and composed of actin filaments and myosin filaments. Z-disc bind the plus ends of actin filaments which extend toward the center of the sarcomere and end at the H-band. Myosin filaments sit in the center of the sarcomere and span the length of the A-band. A-bands are darker in electron micrographs and H&E-stained samples because they contain both actin filaments and myosin filaments. I-bands are lighter because they only contain actin filaments.

Contraction of skeletal muscle is caused by myosin filaments pulling on actin filaments and shortening the distance between the Z-discs. Recall that myosin filaments are bipolar with the motors on opposite sides of the filament trying to move toward opposite Z-discs.
The M-line contains proteins that crosslink adjacent myosin filaments.
Two other important structure seen in this image are T-tubules and sarcoplasmic reticulum. T-tubules are invagination of the cell membrane that allows action potentials on the surface of a muscle cell to penetrate into its center. The sarcoplasmic reticulum is the equivalent of the endoplasmic reticulum in non-muscle cells and serves as a reservoir of calcium.
Types of Skeletal Muscle Cells
Most muscles contain a mixture of these extreme fiber types. In humans, the fiber types cannot be distinguished based on gross examination, but require specific stains or treatments to differentiate the fibers.
Skeletal muscles are divided into two muscle fiber types. Slow-twitch (type I) muscle fibers contract more slowly and rely on aerobic metabolism. They contain large amounts of mitochondria and myoglobin, an oxygen-storage molecule. The reddish color of myoglobin is why these fibers may be referred to as red fibers. These muscles can maintain continuous contraction and are useful in activities such as the maintenance of posture.

Fast-twitch (type II) muscle fibers contract more rapidly due to the presence of a faster myosin. Type II fibers can be subdivided into those that have large amounts of mitochondria and myoglobin and those that have few mitochondria and little myoglobin. The former primarily utilize aerobic respiration to generate energy, whereas the latter rely on glycolysis. The lack of myoglobin results in a paler color than the slow-twitch muscles, and fast-twitch fibers may therefore be referred to as white fibers. These muscles are important for intense but sporadic contractions; for example, those that take place in the biceps.
Please note that the appearance of the type I and type II muscle fibers appear different than what was shown in lecture where the Type I (slow-twitch) appeared light and the Type II (fast-twitch) appeared dark. The difference between the two image is in the method used to stain the samples. Basically, if skeletal muscle is stained under acidic conditions, then the type I fibers stain dark (above image), but if done under alkaline conditions, then type II fibers stain dark (lecture image). For more information please visit HERE
Neuromuscular Junction
Skeletal muscle cells are innervated by motor neurons. A motor unit is defined as the neuron and the skeletal muscle cells it supplies. Some motor neurons innervate one or a few muscle cells whereas other motor neurons can innervate hundreds of muscle cells. Motor axons terminate in a neuromuscular junction on the surface of skeletal muscle fibers.

Neuromuscular Junction Synapse
The neuromuscular junction is composed of a pre-synaptic axon terminal and a post-synaptic muscle cell. Upon depolarization of the axon, synaptic vesicles containing the neurotransmitter acetylcholine fuse with the membrane, releasing acetylcholine into the cleft. Acetylcholine binds to receptors on the post-synaptic membrane and causes depolarization of the muscle fiber, which leads to its contraction. Typically, one action potential in the neuron releases enough neurotransmitter to cause one contraction in the muscle fiber.

Note the basal lamina that surrounds the muscle cell. The basal lamina is similar to the basement membrane of epithelia. The basal lamina of skeletal muscle cells contains acetylcholinesterase which is an enzyme that digests acetylcholine. Acetylcholinesterase helps limit the duration of each contractile event.
Denervation of Skeletal Muscle
Connections between motor neurons and skeletal muscle cells are not only required to trigger contraction of muscle cells, they are also important for maintaining the viability of muscle cells. The viability of skeletal muscle cells depends upon sufficient stimulation by acetylcholine. Reduced exposure to acetylcholine causes skeletal muscle cells to atrophy compromising the strength of the overall muscle tissue.
Cardiac Muscle
Cardiac muscle shares important characteristics with skeletal muscle. Functionally, cardiac muscle produces strong contractions like skeletal muscle, and histologically, cardiac muscle cells appear striated like the skeletal muscle cells due to the presence of sarcomeres.
The cells in cardiac muscle are much shorter than skeletal muscle cells and are arranged in series to span the length of the muscle. Cardiac muscle cells also branch at the ends to form connections with multiple adjacent cells, resulting in a complex, three-dimensional network. In contrast, skeletal muscle cells are arranged into long, parallel arrays.

Cardiac muscle cells contain one to two nuclei and located centrally within the cells. The cells are surrounded by numerous capillaries and contain an abundance of mitochondria to meet the energy demands of the cells.
The cardiac muscle fibers are joined end to end by specialized junctional regions called the intercalated discs. The intercalated discs provide anchorage for myofibrils and allow rapid spread of contractile stimuli between cells. Such rapid spread of contraction allows the cardiac muscles to act as a functional syncytium.
Cardiac Muscle Electron Micrograph
Similar to the cytoplasm of skeletal muscle cells, the cytoplasm of cardiac muscle cells is organized into sarcomeres. Also note the abundance of mitochondria. The dark line is an intercalated disc that connects two adjacent cardiac muscle cells. The intercalated discs contain three types of membrane-to-membrane contacts:
- Adhering junctions that are connected to actin filaments to transmit contraction
- Desmosomes that connect to intermediate filaments and provide mechanical support to the entire tissue
- Gap junctions that allow the spread of current (ions) between adjacent cells and allow cardiac muscle to contract in a coordinated fashion

Smooth Muscle
Smooth muscle is found throughout the body and forms the contractile portion of the walls of the digestive tract from the middle portion of the esophagus to the internal sphincter of the anus; the walls of the respiratory tract from the trachea to the alveolar ducts; the walls of the urinary tract; and the walls of the arteries, veins, and large lymph vessels. Smooth muscle is specialized for slow and sustained contractions of low force.
Smooth muscle fibers are elongated spindle-shaped cells with a single nucleus. The nucleus is located centrally and the sarcoplasm is filled with myosin and actin filaments. Importantly, these filaments are not arranged into sarcomeres as they are in skeletal and cardiac muscle cells.

Smooth Muscle Orientations
The arrangement of smooth muscle differs from organ to organ. Usually, groups of smooth muscle cells will be oriented in one direction to provide contractile force in that direction. This image is a cross section of the ileum. The smooth muscle in the intestine is arranged into two layers. In the layer at the top of the image (layer 1), the smooth muscle cells are arranged to contract in the direction indicated. Note the nuclei in these cells are flat and elongated. The layer at the bottom of the image (layer 2) contains smooth muscle cells that are oriented to contract into and out of the plane of the screen. Note that the nuclei of these cells are round.

Smooth Muscle Cell Electron Micrograph
The thick (myosin) and thin (actin) filaments are scattered throughout the sarcoplasm of smooth muscle cells and are attached to dense bodies on the cell membrane and within the cytoplasm. Since the contractile proteins of these cells are not arranged into myofibrils like those of skeletal and cardiac muscle, they appear smooth rather than striated. Activation of the myosin filaments pulls the dense bodies closer together causing the cell to shrink.

Nervous System
Introduction
Our discussion of the nervous system will focus on the structure and function of the peripheral nerves.
Spinal Cord
Although the spinal cord is part of the central nervous system, many neurons with axons in peripheral nerves have their cell bodies in the spinal cord. In addition, peripheral nerves send projections into the spinal cord. Therefore, an understanding of the architecture of the spinal cord is important for understanding peripheral nerves.
Spinal Cord
Many important features of the spinal cord are visible in this cross section. The white matter is composed of nerve fibers carrying information to and from the brain and makes up the outer regions of the cord. The nerve fibers contain mostly myelinated axons which accounts for the light appearance of the region. The grey matter, which is located in the center of the cord, contains neuronal cell bodies and the axons and dendrites that emanate from those cell bodies. The presence of the cell bodies lends this region a darker color. The grey matter is divided into the dorsal (posterior) horn and ventral (anterior) horn. The ventral horn contains the cell bodies of motor neurons whereas the dorsal horn contains cell bodies of neurons that receive inputs from sensory neurons.

Attached to the spinal cord are two prominent structures: the dorsal root and ventral root. The ventral root contains the axons of motor neurons whose cell bodies reside in the ventral horn matter.
Dorsal Root
The dorsal root contains the cell bodies of sensory neurons that bring information from the periphery to the spinal cord. These neurons are pseudounipolar and contain an axon-like process that bifurcates with one branch extending toward the periphery and the other branch heading toward the grey matter of the spinal cord. Fibers heading toward the periphery leave the ganglion through the spinal nerve, where they run together with motor fibers. The dorsal root ganglion also contains satellite cells, which provide structural and metabolic support to the sensory neurons.

Motor Neurons
Motor neurons innervate one or many muscle fibers to control muscle contraction. Motor neurons are typically multipolar with an axon that terminates in a neuromuscular junction on the surface of skeletal muscle fibers. The dendrites of motor neurons are innervated by neurons in upper regions of the spinal cord.
The motor neuron in the ventral horn is easily identifiable by its large size, polygonal shape and extension from the cell body. Compare the size of the nucleolus in the motor neuron with the nuclei in the surrounding support cells.

Another useful stain for neurons is Nissl which labels rough endoplasmic reticulum in neurons. The dark blue structures are referred to as Nissl bodies but are the equivalent of the rough endoplasmic reticulum. Note that Nissl bodies are confined to the cell bodies and dendrites; they do not extend into the axon.

Peripheral Nerve Bundle
Similar to skeletal muscle, peripheral nerves contain three layers of connective tissue. At the smallest level are the axon, its myelin sheath, if present, and a thin layer of connective tissue called the endoneurium. Several of these axons are bundled together and surrounded by a second layer of connective tissue called the perineurium. Finally, an outer sheath of epineurium surrounds the entire nerve. The epineurium and perineurium contain blood vessels, whereas the endoneurium lacks vasculature.
Peripheral Myelinated Nerve Bundle
This image shows the three connective tissue layers of peripheral nerve that contains myelinated axons. Note that each axon is surrounded by a white space which is where the myelin sheath resides. Because myelin is mostly lipid, it stains poorly in histological samples. A thin layer of pink-staining connective tissue (endoneurium) is visible around the myelin. The perineurium is a thicker layer of connective tissue that encases several axons. Finally, epineurium is the thickest layer of connective tissue and envelopes the entire nerve.

Myelinated Axon - Electron Micrograph
This electron micrograph shows a cross section of a myelinated axon in the peripheral nervous system. A Schwann cell surrounds the axon and wraps it in a sheath of myelin. Collagen fibers of the endoneurium surround the axon and Schwann cell.

Peripheral Unmyelinated Nerve Bundle
Nerves that carry unmyelinated axons contain the same three layers of connective tissue that are found in nerves with myelinated axons. The nuclei of Schwann cells are visible but the axons are difficult to see because they are not set off against the light staining of the myelin sheath.

Unmyelinated Axons EM
Unmyelinated axons are still surrounded by Schwann cells but are not wrapped in myelin sheaths. This EM image shows a Schwann cell associated with several small axons but without myelinating the axons. In unmyelinated nerves, Schwann cells can associate with several axons; whereas in myelinated nerves, Schwann cells associate with only one axon.

Peripheral Nerve - Longitudinal Section
Viewed longitudinally, peripheral nerves appear to have a wave-like form. This arrangement lends peripheral nerves elasticity and extra length to be stretched when a limb is extended.

Peripheral Ganglion
Some organs contain collections of nerve cell bodies organized into ganglia. For example, the small intestine contains numerous ganglia located in its smooth muscle layers. The dendrites of the neurons in the peripheral ganglia receive impulses from nerves extending from the spinal cord, and the axons of the neurons synapse with cells in the organ to regulate their activity. Besides the cell bodies of neurons, peripheral ganglia also contain numbers satellite or support cells.
