Connective Tissue
Introduction
Connective tissue is a term used to describe the tissue of mesodermal origin that that forms a matrix beneath the epithelial layer and is a connecting or supporting framework for most of the organs of the body.
Overview of Connective Tissue
In contrast to epithelia, connective tissue is sparsely populated by cells and contains an extensive extracellular matrix consisting of protein fibers, glycoproteins, and proteoglycans. The function of this type of tissue is to provide structural and mechanical support for other tissues, and to mediate the exchange of nutrients and waste between the circulation and other tissues. These tissues have two principal components, an extracellular matrix and a variety of support cells. These two components will be the focus of this lab.
Extracellular Matrix Components of Connective Tissues
The protein components of the extracellular matrix of connective tissue can be divided into three classes: collagenous fibers, elastic fibers and glycoproteins. In addition to the protein components, the extracellular matrix also contains hyaluronan which is a long chain of disaccharides.
Collagenous Fibers
Collagenous fibers consist of types I, II, or III collagen and are present in all types of connective tissue. They primarily resist tension. In electron micrographs, type I collagen has characteristic appearance of bundles of fibrils which can be seen in cross-session and longitudinally. Higher magnification of the fibrils reveals a repeated banding pattern which is produced by the spacing of the trimers in the fibril.

Connective tissue is often classified based on the amount and orientation of the collagenous fibers. Dense connective tissue contains a large amount of collagenous fibers that are oriented in the same direction and is found structure that resist tension in one direction, such as tendon. Dense, irregular connective tissue also contains a large number of collagenous fibers but these are oriented in multiple directions. This type of connective tissue is found in structures that need to resist tension in multiple directions, such as skin. Lastly, loose connective contains few collagenous fibers; instead, it contains more glycoproteins and cells. This is the most common type of connective tissue and is found in most organs in the body.

Elastic Fibers
Elastic fibers stretch under tension but generate a recoil force when the tension is relaxed. Elastic fibers are prominent in the walls of large arteries where the allow the wall of the artery to stretch during systole and then generate a recoil during diastole. Elastic fibers are a composite material of the proteins elastin and fibrillin. Under the electron microscope, elastin appears as an amorphous mass lined by barely discernable microfibrils of fibrillin. Elastic fibers are stretchable because they are normally disorganized - stretching these fibers makes them take on an organized structure. When the tension is relaxed, the fibers return to their disorganized state.

Proteoglycans
Proteoglycans are proteins which are covalently attached to long disaccharides. The disaccharide side-chains are linked to proteins in the ER and Golgi. Because the disaccharides are negatively charged, they attract sodium which draws in water to the connective tissue. The retention of water allows connective tissue to resist compression. Because proteoglycans stain poorly by H&E, they are difficult to see in most histological samples.
Reticular Fibers
Reticular fibers are composed of type III collagen. Unlike the thick and coarse collagenous fibers, reticular fibers form a thin reticular network. Such networks are widespread among different tissues and form supporting frameworks in the liver, lymphoid organs, capillary endothelia, and muscle fibers.

Cells of Connective Tissue
Although the connective tissue has a lower density of cells than the other tissues you will study this year, the cells of these tissues are extremely important.
Fibroblasts
Fibroblasts are by far the most common native cell type of connective tissue. The fibroblast synthesizes the collagen, elastic fibers and proteoglycans of the extracellular matrix. These cells make a large amount of protein that they secrete to build the connective tissue layer. Some fibroblasts have a contractile function; these are called myofibroblasts. This electron micrograph shows several fibroblasts. Note the large nuclei, with heterochromatin and euchromatin, as well as the abundant rough endoplasmic reticulum. Fibroblasts can be seen by light microscopy in type of loose connective tissue called areolar tissue.

Mast Cells
Mast cells are granulated cells typically found in connective tissue. These cells mediate immune responses to foreign particles. In particular, they release large amounts of histamine and enzymes in response to antigen recognition. This degranulation process is protective when foreign organisms invade the body, but is also the cause of many allergic reactions. Mast cells can be identified in connective tissue by their numerous cytoplasmic granules.

Adipocytes
Adipocytes or white fat cells are specialized for the storage of triglyceride, and occur singly or in small groups scattered throughout the loose connective tissue. They are especially common along smaller blood vessels. When fat cells have accumulated in such abundance that they crowd out or replace cellular and fibrous elements, the accumulation is termed adipose tissue. These cells can grow up to 100 µm and usually contain once centrally located vacuole of lipid - the cytoplasm forms a circular ring around this vacuole, and the nucleus is compressed and displaced to the side. The function of white fat is to serve as an energy source and thermal insulator.

Brown Fat Cells
Brown fat cells are highly specialized for temperature regulation. These cells are abundant in newborns and hibernating mammals, but are rare in adults. They have numerous, smaller lipid droplets and a large number of mitochondria, whose cytochromes impart the brown color of the tissue. The electron transport chain of these mitochondria is disrupted by an uncoupling protein, which causes the dissipation of the mitochondrial hydrogen ion gradient without ATP production. This generates heat.

Macrophage
Macrophages are phagocytic cells that are capable of engulfing foreign antigens and remnants of dead cells. Macrophages descend from monocytes and are found in most organs where they have different names depending on the organ. This image shows a macrophage with an irregularly shaped nucleus. The cytoplasm of the macrophage contains phagosomes and residual bodies, which are lysosomes with undigested material. On the right is a macrophage in the airway in the lung. These macrophages are called dust cells.

Cartilage
Cartilage is a specialized form of connective tissue produced by differentiated fibroblast-like cells called chondrocytes. It is characterized by a prominent extracellular matrix consisting of various proportions of connective tissue fibers embedded in a gel-like matrix rich in glycoproteins and hyaluronan. Chondrocytes produce all of the structural components of cartilage, including collagen, proteoglycans and glycosaminoglycans. Note the basophilia of the cytoplasm and the presence of lipid droplets.

Three kinds of cartilage are classified according to the abundance of certain fibers and the characteristics of their matrix.
Hyaline Cartilage
Hyaline cartilage is the most common type of cartilage and has a matrix composed of type II collagen and chondromucoprotein, a copolymer of chondroitin sulfates A and C (a disaccharide) with protein. Its high concentration of negatively-charged sulfate groups makes it appear intensely basophilic under H&E, and it often has a glassy appearance. Note the numerous chondrocytes in this image, surrounded by the cartilage they have produced. These cells have relatively small nuclei and often demonstrate lipid droplets in their cytoplasm. The spindle-shaped cells in the perichondrium can differentiate into chondroblasts that will eventually develop into chondrocytes. This cartilage is found in the nose, tracheal rings, and where the ribs join the sternum.

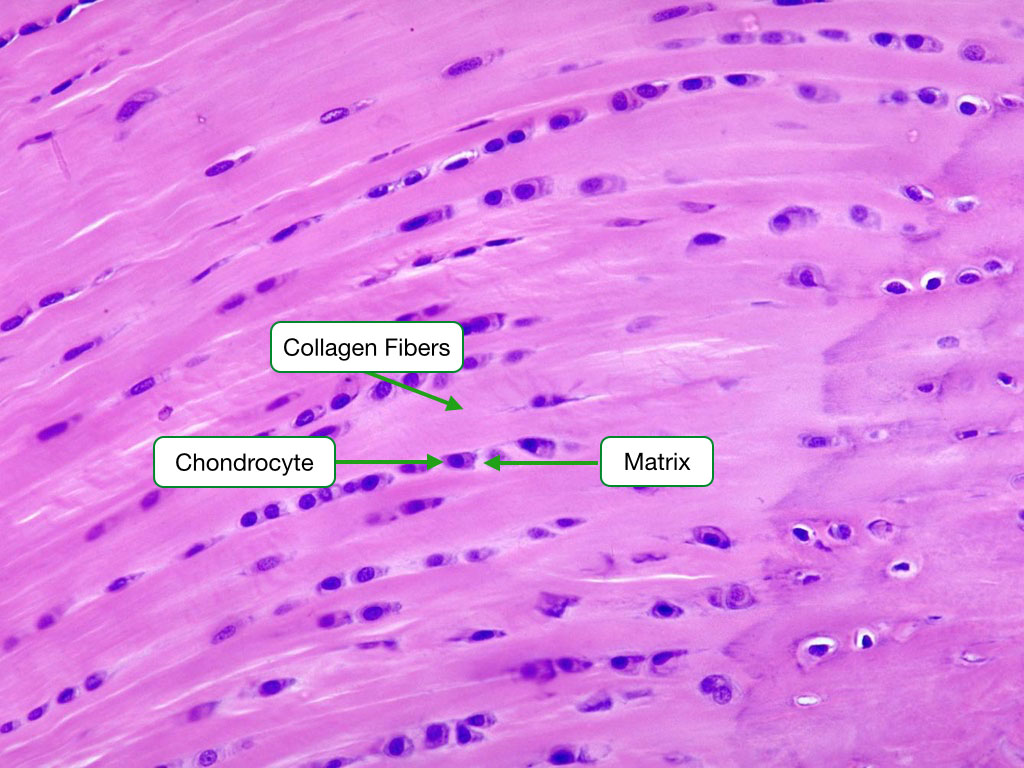
Fibrocartilage
Fibrocartilage is distinguished by its high content and orderly arrangement of type I collagen fibers. It is typically located in regions where tendons attach to bones, the intervertebral discs, and the pubic symphysis. Numerous chondrocytes are spaced between the fibers. Note that the chondrocytes are surrounded by a matrix which helps differentiate fibrocartilage from dense connective tissue.
Elastic Cartilage
Elastic cartilage is characterized by the presence of abundant elastic fibers and is quite cellular. It is made up of type II collagen and is located in the auricle of the ear and the epiglottis.

Bone
Introduction
Bone is a form of connective tissues with special properties and serves several functions:
- Mechanical support for sites of muscle attachment
- Protection of vital organs
- Reservoir for calcium and phosphate
- Site for development of blood cells
Macroscopic Organization of Bone
Bone comes in two general architectures. Compact bone which composes the outer wall of most bones and trabecular bone which is found in the inner cavities of bone. A long bone illustrates both types of bone. Trabecular bone is found in the head region or epiphysis of the bone and appears as many struts or spicules of bone. A thick layer of compact bone is found along the shaft or diaphysis of the long bone.

Long bone also illustrates the two surface of bone. The periosteal or outer surface of the bone is normally covered with a layer of fibrous tissue called periosteum and is the site where muscle and tendon attaches to bone. The inner surface bone is called the endosteal surface and is usually covered by a thin layer of cells called the endosteum.
All bone is composed of type I collagen fibrils that are encased in crystals of calcium-phosphate.
Trabecular Bone
This image illustrates trabecular bone. Note the thin, branched spicules of bone. Within the bone matrix are osteocytes. Around the trbeculae is bone marrow.

Compact Bone
Compact bone is organized into parallel columns, known as Haversian systems, which run lengthwise down the axis of long bones. This image shows compact bone in cross section. Haversian systems comprise concentric rings of bone around a central channel or Haversian canal. The canal contains nerves, blood vessels, and lymphatic system of the bone. Also seen in this image is a Volkmann's canal. These canals start at the periosteal surface and contain blood vessels that connect to blood vessels in the Haversian canals. This image and the next are an example of unstained bone.

Haversian System
Osteoblasts synthesize the concentric rings of bone in the Haversian systems. As osteoblasts secrete matrix, they become trapped in spaces called lacunae and become known as osteocytes. Osteocytes communicate with each other and with the Haversian canal through cytoplasmic extensions called filopodia that run through small interconnecting canals in the bone called canaliculi. Filopodia from adjacent osteocytes communicate via gap junctions.

Osteoblasts
Osteoblasts synthesize new bone by secreting collagen fibrils and facilitating the mineralization of the fibrils. Active osteoblasts sit atop a layer of osteoid that contains collagen fibrils that have not completely mineralized. Over time the fibrils in the osteoid with mineralize to form bone.

Osteoclasts
Osteoclasts are large, multinucleated cells that derive from monocytes and therefore are related to macrophages. They resorb bone by secreting organic acids, which dissolve hydroxyapatite, and enzymes, which digest the collagen fibrils. Active osteoclasts create surface depressions in bone due to reposition of the bone.

Activation of Osteoclasts
Recall from lecture that osteoclasts derive from monocytes that differentiate into pre-osteoclasts. Pre-osteoclasts transform into osteoclasts when they bind RANK ligand on the surface of osteoblasts. When the body needs more calcium, it produces parathyroid hormone which increases the the number of osteoclasts.

Bone Modeling and Remodeling
Osteoclasts and osteoblasts often work together to reshape bone (modeling) and replace old bone with new bone (remodeling). In bone modeling the osteoclasts and osteoblasts work on opposite surfaces of bone to position the bone in one direction. In bone remodeling, osteoclasts and osteoblasts work on the same surface of bone to digest old bone and replace it with new bone. Proper balance between the number and activity of osteoblasts and osteoclasts is essential to ensure that bone is neither overproduced nor over degraded.

Development of Bone
During development, bone can generally only form on a preexisting structures. The two most common structures are mesenchymal tissue and cartilage. Bone forming on mesenchyme is called intramembraneous ossification, whereas bone forming on cartilage is called endochondrial ossification. These are important not only during development, but the same mechanisms are used to repair bone when it fractures.
Intramembraneous Ossification
Intramembranous ossification is the direct conversion of embryological mesenchymal tissue to bone. The process begins when mesencyhmal cells differentiate into osteoblasts, which begin to synthesize osteoid that will eventually mineralize into bone.

Endochondrial Ossification
In endochondrial ossification, bone is synthesized over a cartilage template. This image shows a growing tibia. The purple growth plate is composed of cartilage synthesized by the embedded chondrocytes. Over time, the cartilage becomes calcified; the dark purple areas within the trabecuale are remnants of calcified cartilage. As the cartilage emerges from the growth plate, woven bone, which appears light blue in this slide, is laid down over the calcified cartilage. This preliminary bone will eventually be replaced through bone remodeling to produce more organized lamellar bone. Osteoid, appearing red, can be seen laid down over the primary trabeculae of woven bone with a cartilage core.

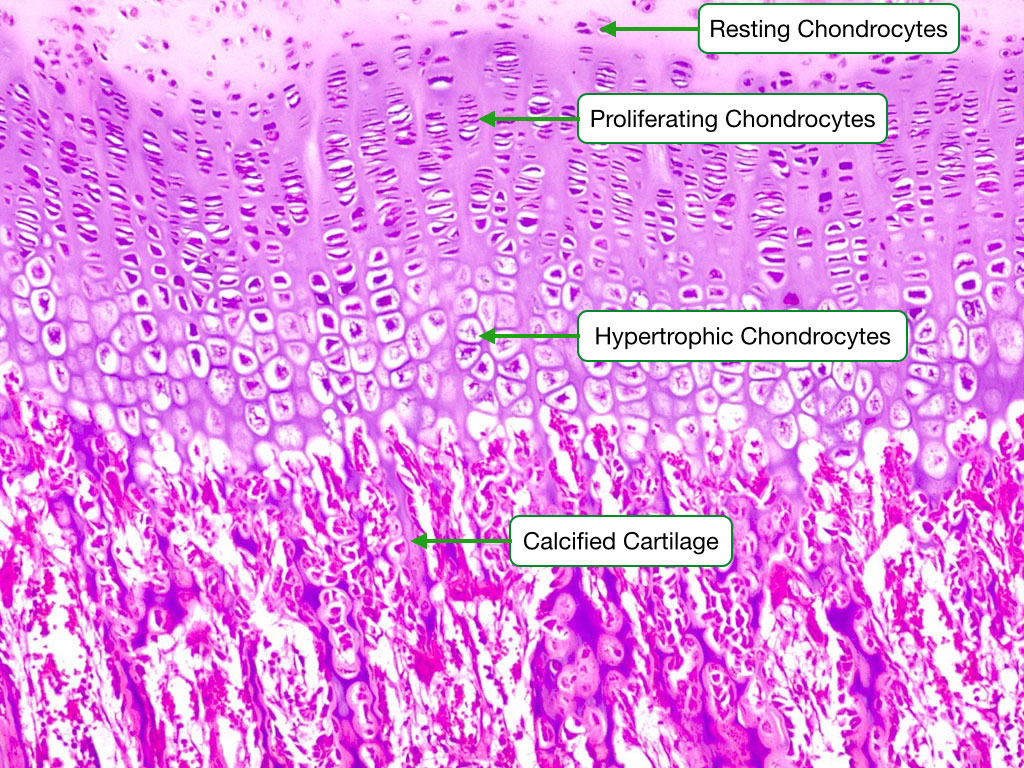
Chondrocyte Growth Sequence
This image shows the different stages of chondrocyte development during endochondrial ossification. In the initial stage, the chondrocytes are resting and the cartilage is not being converted to bone matrix. They then go through a stage of proliferation, where each lacuna contains numerous chondrocytes. This is followed by maturation/hypertrophy phases in which the chondrocytes and their lacunae appear much larger than when they were resting. The cartilage is eventually calcified. Finally, the calcified cartilage is adsorbed by osteoclasts, woven bone is laid down and after ~70 days is converted to lamellar bone.