Immune System
An immune response is divided into two processes: innate and adaptive. An innate response is a rapid but non-specific reaction to foreign pathogens. An adaptive response is slower to develop but generates a reaction against a specific pathogen and importantly, a memory of that pathogen that is stored for to fight future infections by the pathogen. An adaptive response has two branches: cell-mediated and humoral. A humoral immune response generates antibodies against specific molecules on a foreign pathogen. This TBL will focus on the structures that facilitate the development of a humoral immune response.
A humoral immune response requires the activation of a small number of B-cells that are capable of producing antibodies that bind to regions of macromolecules (antigens) on a pathogen. To activate a B-cell, its receptor (B-cell receptor) must bind with sufficient affinity to an antigen. The B-cell takes up its B-cell receptor bound to antigen via endocytosis and then presents a portion of the antigen on its surface in complex with class II MHC. The B-cell then waits to be directly stimulated by an activated T-cell. A T-cell is activated when its receptor (T-cell receptor) binds with sufficient affinity to the antigen presented in a complex with class II MHC on the surface of an antigen-presenting cell (APC). The interaction between the T-cell and APC activate the T-cell. The T-cell is now capable to activate the B-cell that presents the same antigen on its surface complexed with class II MHC.

These interactions and how they lead to activate B and T cells have been well-studied and can be readily recapitulated in vitro. However, the immune system faces an enormous challenge in vivo because only a tiny fraction of all the B or T cells in the body recognize a specific antigen. These cells are distributed throughout the body and the probability that any of these cells encounters an antigen during infection would seem unlikely. This TBL will explore the structure and function of two organs that facilitate immune responses: lymph nodes and the spleen. In addition, the lab will examine the thymus where T-cells mature.
Lymph Node
Lymph nodes facilitate the interaction between antigens, antigen presenting cells, and lymphocytes to generate humoral immune response. Lymph nodes occur along the course of the lymphatic vessels that drain lymph from organs and tissues. They filter the lymph before it drains back to the bloodstream and are therefore, in an ideal position to collect antigens from local infections. Normally, they are only a few millimeters in diameter. However, when an immune response is initiated against antigens, the lymphocytes within the lymph nodes undergo activation and proliferation, causing the nodes to enlarge.
Cross-section of Lymph Node
Lymph nodes are usually bean-shaped, with an indented region known as the hilum. They are covered by a collagenous capsule that extends into the body of the node as trabeculae. The body of the lymph node is divided into an outer cortex and an inner medulla. The cortex contains a high concentration of lymphocytes while the inner medulla is less cellular. The lymphocytes in the cortex self-organized into secondary follicles in response to foreign antigen. Follicles are where B-cells mature and proliferate. A paracortex is also defined in lymph nodes as a site where T-cells accumulate


Flow of Lymph
Lymph from the extracellular space carries antigens and antigen presenting cells, such as dendritic cells, from the tissues to the lymph nodes. The lymph enters the node at several points along the lymphatic system through afferent lymphatic vessels. These vessels pierce through the capsule and drain into the space below, known as the sub-capsular sinus. From the sub-capsular sinus, the lymph drains toward the medulla via channels called cortical sinuses. The sinuses are lined by endothelial cells. After reaching the medulla, the lymph drains into a complex network of medullary sinuses. The medullary sinuses converge at the hilum and drain into the efferent lymphatic vessels.

Sampling of Antigen
Macrophages in the parenchyma of the lymph nodes sit underneath the sinuses and extend cellular processes into the sinus channels. These processes capture antigen which is then brought into the parenchyma of the lymph node where it can be sampled by lymphocytes.

Capusle and Subcapsular Sinus
This is a high power view of the lymph node capsule and sub capsular sinus. Note the afferent lymphatic vessels traversing the capsule. The lymphatic vessels contain valves, which are clearly seen in this slide. The sub capsular sinus is lined by a layer of endothelial cells. Beneath the endothelial cells are macrophages that retrieve antigen from the lymph in the sub capsular sinus. These macrophages cannot be distinguished in histological images.

Lymphoid Follicles
In the cortex, B-lymphocytes are localized in lymphoid follicles just beneath the capsule. In absence of an active immune response, these follicles are known as primary lymphoid follicles and are difficult to distinguish histologically. When an immune response is underway, the follicles develop germinal centers that contain proliferating and maturing B-cells that are responding to antigen. Surrounding the germinal centers is the mantle zone that contains resting and memory B-cells. T-cells also reside close to germinal centers and help select and stimulate B-cells. Follicles with germinal centers are called secondary lymphoid follicles.

Several important events occur in lymphoid follicles. First, immature and mature B-cells expand in numbers through rapid cell division. Second, the immune response to antigen evolves as the affinities of antibodies synthesized by B-cells increase over time. The increase in affinity is driven by rearrangement and mutation of DNA sequences that encode the antigen-binding segment of antibodies. Antigen is presented to B-cells in germinal centers and those B-cells that express antibodies with high affinity to antigen are selected and stimulated to grow and divide.
Proliferation and selection of B-cells appear to occur in separate zones within the germinal center. Histologically, germinal centers are divided into a dark zone and light zone. The dark zone contains B-cells undergoing cell division which expands the population of B-cells. In the light zone, B-cells encounter antigen presented by follicular dendritic cells. Follicular dendritic cells (FDC) bind and present antigen captured from lymph which percolates through the lymph node. B-cells that express a receptor with high affinity to the presented antigen bind to the surface of FDCs and receive stimulatory signals. Many of these B-cells will receive additional stimulation from T-cells that recognize the same antigen.
In time-lapse videos, B-cells are seen migrating between the two zones. Immature B-cells or centroblasts proliferate in the dark zone and then move to the light zone. If their B-cell receptor binds antigen presented by FDCs and they find their cognate T-cells, they mature to become centrocytes and then migrate back to the dark zone where they divide to expand the number of B-cells that recognize antigen. The subtle difference in staining intensity between the zones is due to the higher concentration of B-cells in the dark zone compared to the light zone which has many light-staining follicular dendritic cells.

Secondary follicles with germinal centers are not unique to lymph nodes but can form anywhere an immune response is generated. They are prominent along the wall of the intestine and in the spleen (see below) but can also form in other organs during an immune response to an infection. When occurring outside the lymph node these follicles are usually called secondary lymphoid organs.
Paracortex
The paracortex lies between the cortex and medulla and contains a high concentration of T-cells. Another feature of this region is the high endothelial venule, where circulating lymphocytes leave the bloodstream to enter the lymph node. High endothelial venues can be distinguished by their cuboidal endothelial cells. Adhesion molecules such as selectins and integrins mediate attachment of lymphocytes to endothelial cells. In a T cell-dominant immunological response, one may observe expansion of the paracortical region.

Medulla
The medulla contains aggregates of lymphoid tissue called medullary cords. The cords contain macrophages and antibody-secreting B-cells called plasma cells. Lymph flows past the cords through lymphatic channels called medullary sinuses. The medullary sinuses merge into efferent lymphatic vessels, which carry the lymph away from the node.

Spleen
The spleen is an organ located in the upper left quadrant of the abdomen. It serves three main functions:
- Activation of immunity against antigens from the blood
- Disposal of old or malformed blood cells, particularly erythrocytes
- Hemopoiesis in fetus (and in adults with particular diseases)
The spleen is covered by an outer capsule that extends into the parenchyma as trabeculae. The majority of the spleen is composed of a matrix called the red pulp, which is the site of erythrocyte disposal. Embedded within the red pulp are small nodules called the white pulp. These nodules contain the lymphocytes.

Blood Flow in the Spleen
The structure of the red and white pulp can be best appreciated by considering the organization of red and white pulp in relation to the blood vessels of the spleen.
The splenic artery enters the spleen at the hillus and branches into trabecular arteries. These arteries exit the trabeculae and split into central arteries, which run through the center of the white pulp.
In the white pulp, the central artery is surrounded by the periarteriolar lymphoid sheath (PALS), which is made up of mostly T-cells. Along the border of the PALS lies the follicle, which consists mainly of B-cells. These follicles can develop into germinal centers, similar to those seen in the lymph node, when exposed to a reactive antigen.
Blood can take two routes to traverse the red pulp. In the closed route (not shown in the image below), blood flows through capillaries and then immediately into the splenic veins. In the open route, blood flows through capillaries and then into the splenic cords which are composed of reticular fibers, reticular cells (a type of fibroblast) and macrophages. To reenter the circulatory system, red blood cells must squeeze through the discontinuous endothelial lining of the venous sinusoids which surround the splenic cords. Old red blood cells that have become less flexible fail to enter the sinusoids and are phagocytosed by macrophages. The splenic sinuses will eventually merge with the splenic vein.

White Pulp
This section of the white pulp shows a central artery surrounded by peri-arteriolar lymphatic sheath (PALS), which is a T-cell zone. It is common to find B-cell lymphoid follicles closely associated with PALSs. The follicle here is a secondary follicle, as evidenced by the presence of a pale-staining germinal center. The germinal center is surrounded by a narrow ring of deep-stained mantle that contains resting B-cells. The mantle separates the follicle from the red pulp.

Red Pulp
The red pulp is mainly filled with venous sinuses, which contain red blood cells and occasionally white blood cells. The sinuses are lined by elongated, rod-shaped endothelial cells that form a discontinuous endothelium. Slits between the endothelial cells allow viable red blood cells, which have entered red pulp through the open route, to squeeze into the sinuses from the splenic parenchyma. These sinuses eventually drain into the splenic vein. Surrounding the sinuses is the parenchyma, which contains macrophages and other immune cells. Macrophages engulf red blood cells that fail to cross the endothelium lining the sinuses.

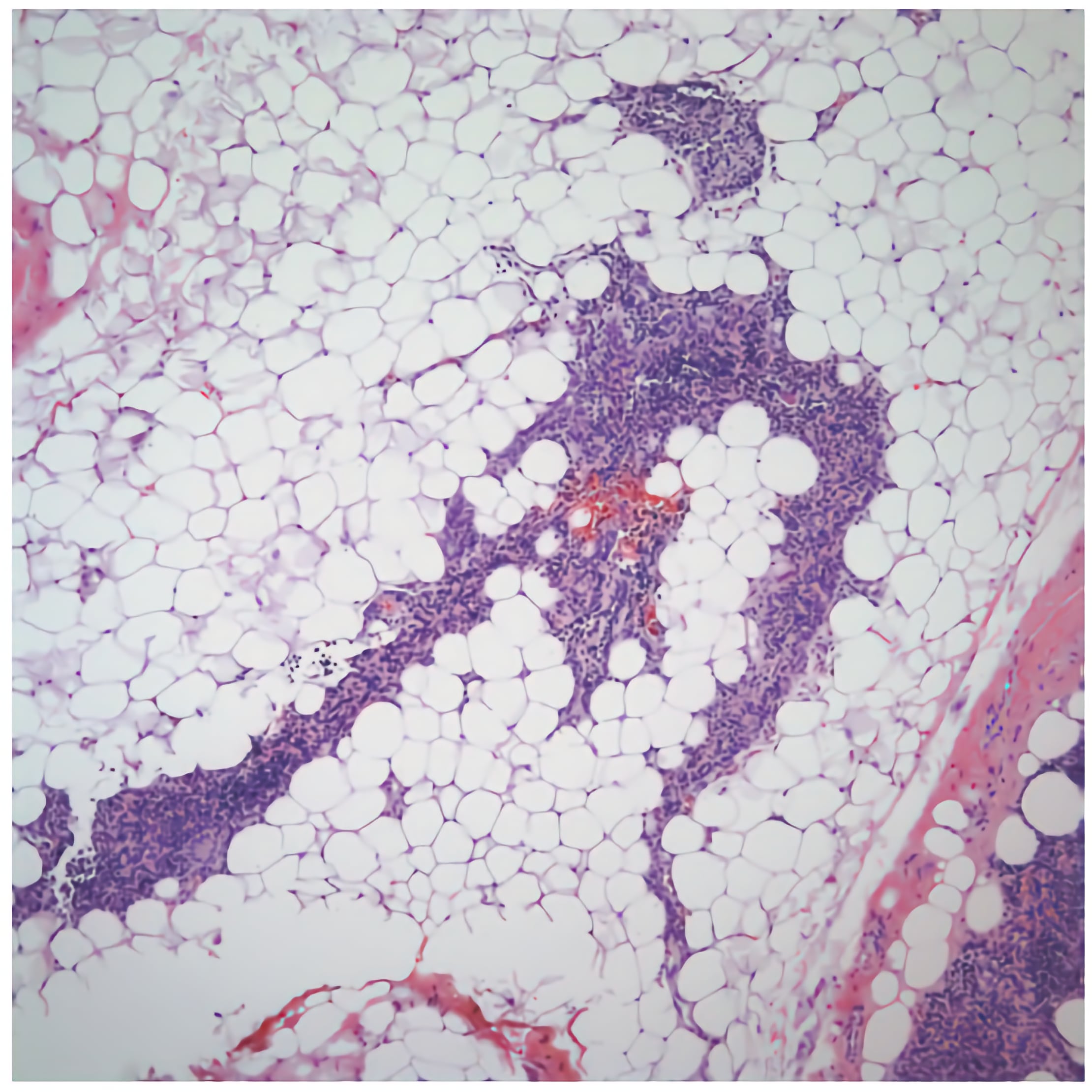
Thymus
The thymus is the primary lymphoid organ engaged in the maturation of T-cells. It is most active during childhood and slowly atrophies after puberty, filling with adipose tissue. Two key events in T-cell maturation occur in the thymus. First is positive selection in which T-cells are selected based on their ability to bind class I or class II MHC. Second is negative selection which eliminates those T-cells whose receptor recognizes self-antigen.
This is a low power view of a young thymus. Note that the gland is organized into numerous lobules. Each lobule features an outer cortex that is densely populated by lymphocytes and an inner medulla that has fewer lymphocytes (and thus is less heavily stained). Also note the loose collagenous capsule that extends into the thymus to form the interlobular septa that separate the lobules. The capsule and septa contain blood vessels, lymphatics and nerves.

Thymus Lobule
T-cells mature in distinct regions of the thymus. Progenitor T-cells, called thymocytes, enter the thymus through blood vessels in the border between the cortex and medulla. The thymocytes migrate to the cortex where they begin the maturation process. As the cells mature, they migrate toward the medulla. Immunocompetent T-cells exit the thymus via post-capillary venules or efferent lymphatics in the medulla.

Thymus Cortex
The thymus cortex contains immature thymocytes and thymocytes undergoing maturation. The immature thymocytes are larger and found in the outer cortex; these cells actively divide and several mitotic cells can be seen in the cortex. Maturation of thymocytes in the cortex starts with the expression of specific cell surface protein (e.g. CD4, CD8 and T-cell receptor). Maturing thymocytes are found deeper in the cortex.

Maturation of thymocytes occurs in two stages: positive selection and negative selection. Positive selection identifies those thymocytes with a T-cell receptor that binds with sufficient affinity to either class I or class II MHC. The cortex contains support cells called cortical epithelial cells that play a critical role in positive selection of T-cells. They express on their cell surface class I or class II major histocompatibility complex (MHC) bound to self-peptide. Only those maturing T-cells with a T-cell receptor that binds with sufficient affinity to the MHC-peptide complex will survive; the rest will undergo apoptosis. Macrophages in the cortex phagocytose apoptotic cells. The cortical epithelial cells can be recognized by their large, pale-staining nuclei.
Thymus Medulla
Negative selection eliminates those thymocytes with a T-cell receptor that binds with high affinity to self-peptides. Negative selection occurs mostly in the medulla and base of the cortex. The medulla contains fewer T-cells and appears more lightly stained than the cortex. The epithelial cells are more easily seen in the medulla. In addition to T-cells and epithelial cells, the medulla contains large numbers of macrophages and dendritic cells. Epithelial cells and dendritic cells play a critical role in negative selection of T-cells. The cells are capable of expressing tissue-specific genes to generate proteins that will be found throughout the body. The cells display peptides from these proteins complexed with MHC on their cell surface. T-cells with receptors that bind strongly to MHC:self-peptide are triggered to undergo apoptosis.

One characteristic feature of the thymus medulla are Hassall corpuscles. These structures are swirls of epithelial cells that can often contain keratin. The function of Hassall corpuscles is currently unknown but they appear to be benign.
Involution of the Thymus
The thymus becomes active in neonates and remains active throughout childhood until puberty when the thymus begins to involute. During involution, the number of thymocytes decreases and production of T-cells slows. Adipose tissue replaces thymocytes until most of the thymus is composed of adipose tissue.