Histology of Cells and Epithelia
Introduction
Histology is the study of the structures of cells and tissues. As you learn about the morphology of cells and the architecture of tissues, you will also learn how those structure indicate and facilitate the functions of cells and tissues. Histology focuses on the normal, how cells and tissues should appear in the absence of disease or infection. Understanding the normal will allow you to identify changes that occur to cells and tissues during disease. To help you with this transition, we will introduce some histological images of diseased tissue.
Preparation of histological samples
Histology involves careful examination of biological samples usually in a light microscope but occasionally in an electron microscope. Preparing histological samples requires several steps that determine which structures and features will be seen in the microscope.
Obtaining a sample
Most samples are usually obtained by biopsy from a patient during surgery or who is anesthetized. Samples are processed quickly to minimize degradation of the specimen.
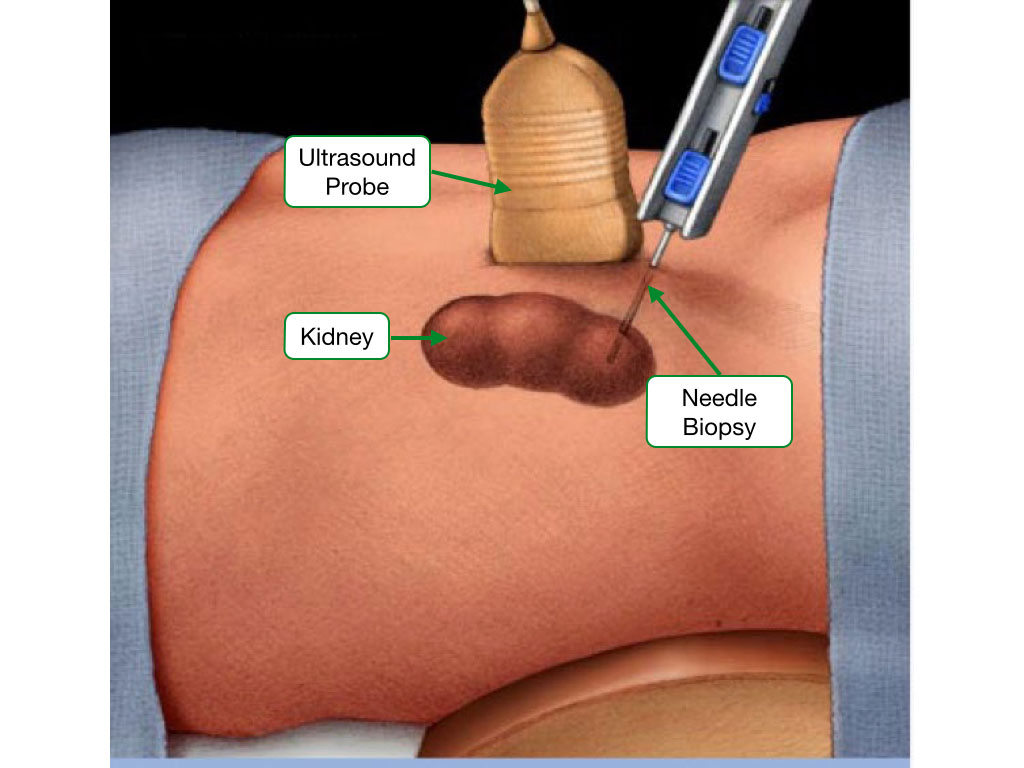
Fixation of the sample
Once removed from the body, all specimens will degrade due to the presence of numerous digestive enzymes in the specimen. To preserve the structure of the specimen, a fixative is applied to inactivate proteins and maintain the integrity of the cells and tissues.
The most common fixative is formaldehyde, also called formalin. Formaldehyde forms chemical cross links between proteins and between nucleic acids. The cross links inactivate most enzymes and preserve the structure that contain protein or nucleic acids. Unfortunately, lipids are not cross linked by formaldehyde and several steps in the processing tissues involve washing with alcohols which solubilizes the lipids are removes them from the sample.

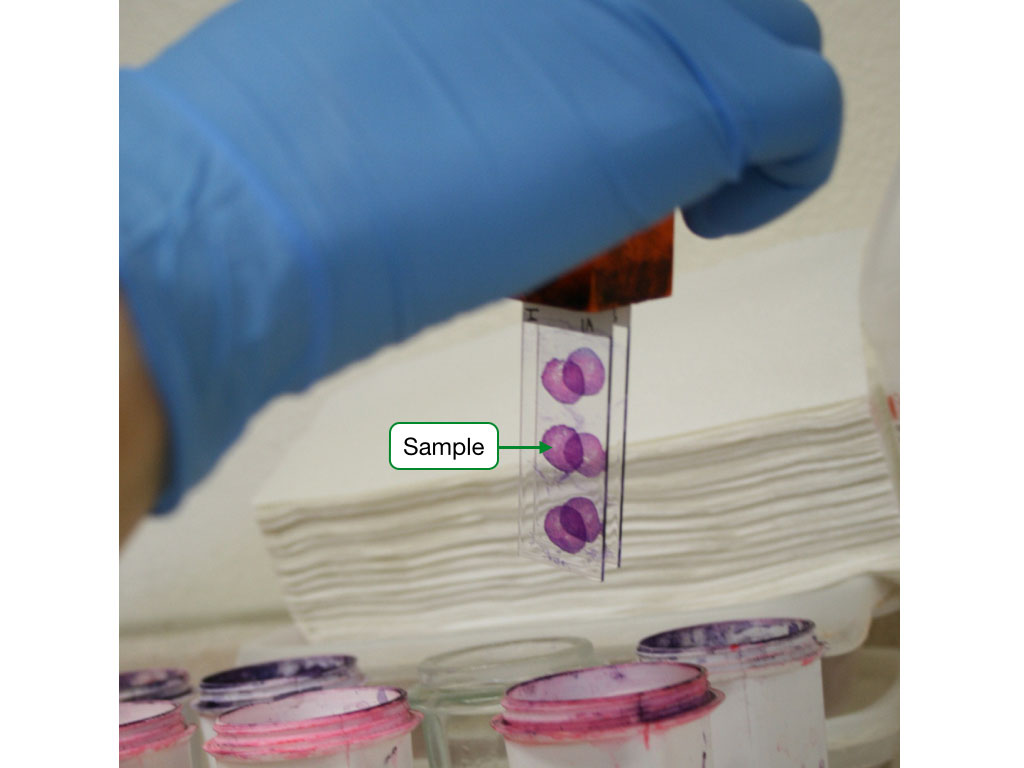
Sectioning the sample
To view a sample under a light microscope, it must be sectioned into very this sliced (about 5 microns). Thin slices produce better imaging of the sample in a microscope, but thinly slicing most samples is difficult due to the softness of the sample. Therefore, most samples are embedded in waxy substance such as paraffin which hardens and allows the sample to be sliced into thin sections.

Staining the sample
Most biological specimens offer poor visibility under standard light microscopy. To increase visibility and help us identify specific structures, histological samples are stained with specific dyes. The dyes associate with specific chemical features of the sample and impart a color. By using combinations of dyes we can label different structures in histological samples.
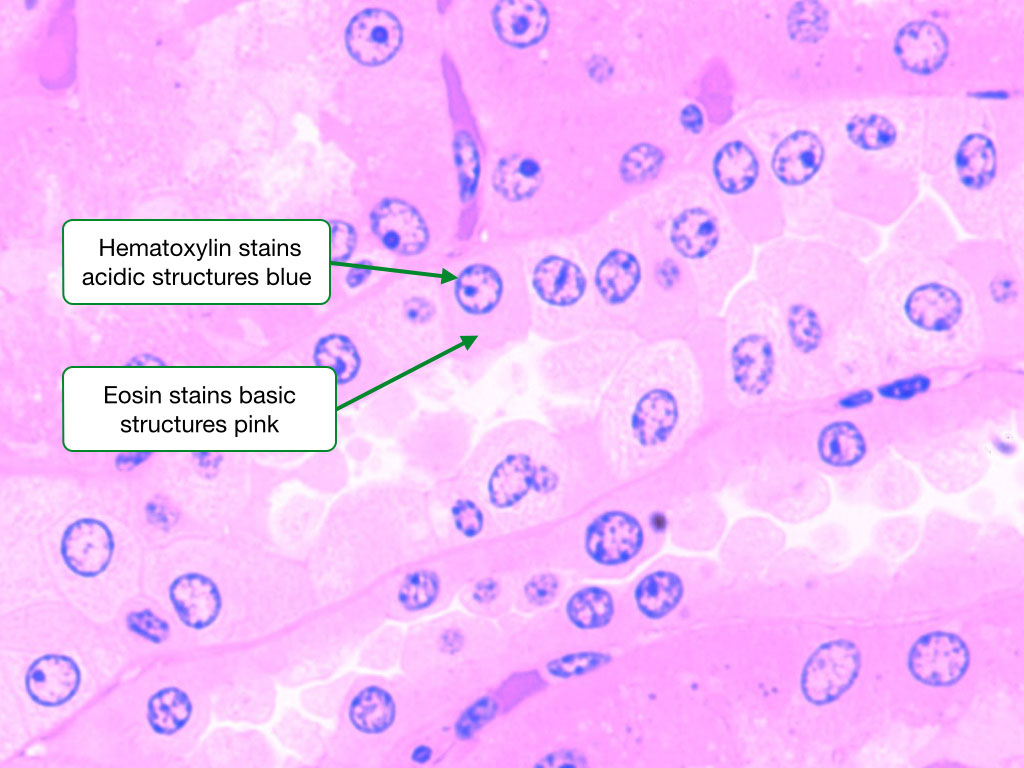
H&E Stain
By far, hematoxylin and eosin (H&E) is the most commonly used stain in histology and pathology. Hematoxylin binds negatively charged structures in samples and imparts a blue or purple color to those structures. The largest and most negatively charged structures in cells are nucleic acids such as DNA in chromosomes and RNA in ribosomes. Hematoxylin consequently strongly labels the nucleus and rough endoplasmic reticulum.
In contrast, eosin labels structures with a positive charge and imparts a pinkish color. There are few large structures with strong positive charge in cells, so eosin does not distinctly label all cells as does hematoxylin. Eosin gives the cytoplasm of most cells a pinkish hue and the secretory granules in some cells are labeled by eosin.
Other stains
There are several additional stains that are used for histological samples. For example, periodic acid–Schiff stain (PAS) labels structure with a lot of carbohydrate (e.g. extracellular matrix, mucus), and is used to diagnose certain diseases, such as glycogen storage diseases. Another common stain is Masson’s trichrome which is often used to label connective tissue. Most of the samples used in the subsequent labs will have been stained with H&E, but samples labeled with a stain will be indicated.
What you can see in a histological image
Histological images often resemble modern art with seemingly random swirls of color and no apparent point of focus. However, in most images there are several identifiable structures that can help orient you in the image and make sense of what you are looking at.
Nucleus
Because nuclei of cells are usually strongly labeled by hematoxylin and appear deep blue, they are often the easiest cellular structure to identify. In addition, much can be learned about the identity, function and activity of a cell from the shape and staining pattern of its nucleus.

Nucleus morphology and location
The shape of the nucleus can he helpful for identifying a cell, especially in blood smears, or finding areas of pathology. Although most cells have nuclei that are round or oval, some cells have nuclei with very distinct shapes. Some white blood cells have nuclei that are divided into lobes and the cells can be identified based on the number of lobes. In addition, some cells have nuclei in specific locations within the cell.

Transcriptional activity
All nuclei contain DNA arranged in chromosomes, but the arrangement of the chromosomes within the nucleus can often indicate how actively a cell is transcribing its genes into mRNA. Structurally, chromosomes are often divided into heterochromatin and euchromatin. In heterochromatin, the chromosomes are condensed and often localize to the edge of the nucleus called the nuclear envelope. Genes in heterochromatin are usually transcriptionally silent. In euchromatin, the chromosomes are less condensed, tend to be located more centrally within the nucleus and have more genes being transcribed.

Dividing cells
Actively dividing cells can be identified by looking at the structure of the nucleus and arrangement of chromosomes. When cells go through mitosis the nuclear envelope breakdown and the chromosomes condense. Eventually, the chromosomes are corralled onto the spindle and then segregated into the two daughter cells. All of these events are visible in histological samples stained by H&E. Identifying and counting mitotic cells in a tissue is often used to determine whether a tissue has become cancerous.

Number of cells
Most cells have one nucleus and therefore, the number of cells in a tissue can be estimated by counting the number of nuclei. Often a change in the number of nuclei can indicate disease. If a sample contains more nuclei (cells) than expected, it can indicate inflammation due to an influx of immune cells or cancer due to over proliferation of cells in the sample. In contrast, if a sample contains fewer nuclei (cells) than expected, it can indicate tissue damage and cell death. Becoming familiar with the relative number of cells in the different tissues and organs is important for identifying and diagnosing disease in histological samples.

Nucleolus
Within the nucleus lies the nucleolus which can also be seen in histological samples. The nucleolus is the site where ribosomes assemble and contains a high concentration or ribosomal genes and RNA. The nucleolus often localizes to the center of the nucleus and stains deep blue. Cells that are actively synthesizing protein require more ribosomes and consequently, the nucleolus in those cells is more prominent. Thus, a prominent nucleolus indicates a cell that is synthesizing a lot of protein.

Rough endoplasmic reticulum
Within the cytoplasm the rough endoplasmic reticulum (ER) is readily seen in cells that are secreting large amount of protein. Because the rough ER contains a high concentration of ribosomes which contain RNA, it will stain blue in H&E samples and contrast nicely with the pinkish cytoplasm.

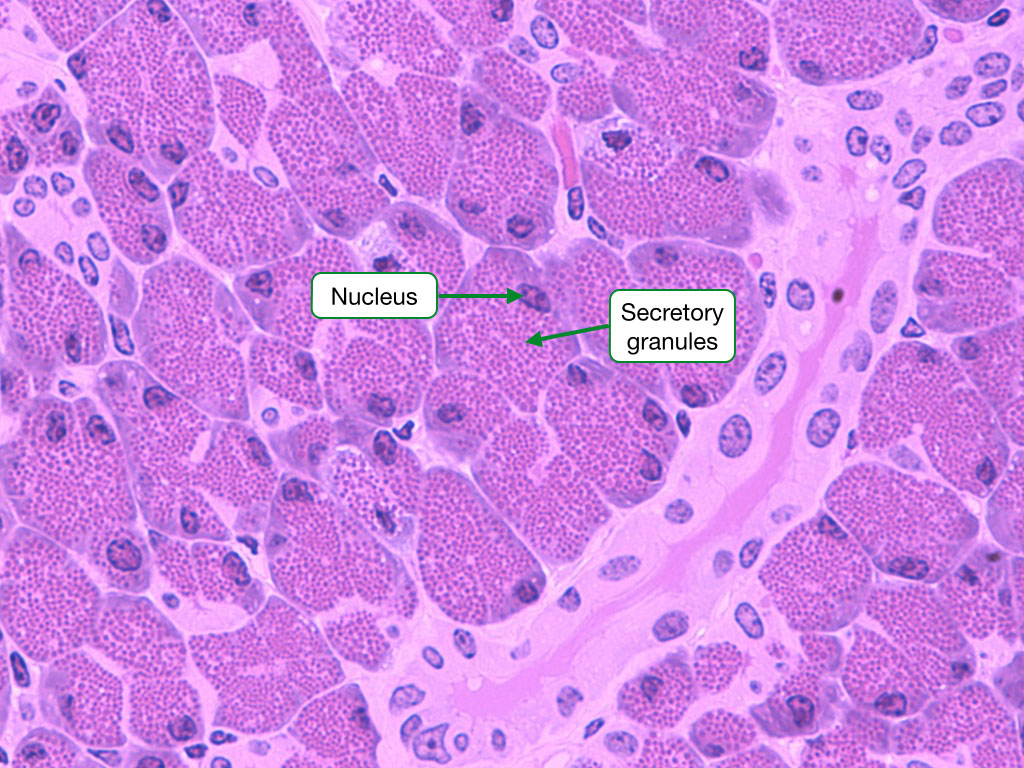
Secretory granules
Some secretory cells store their products in vesicles within their cytoplasm. These vesicles are called granules and when stimulated, the cell will allow the granules to fuse with the cell membrane, releasing the contents of the granules to the surrounding environment. Secretory granules are often visible in H&E samples and their color varies depending on the type of cell.
Estimating the size of cells and other structures
In many histological images, the authors will include a small line of specified length that can be used as a ruler to measure the size of cells and other objects. There is no ruler, however, when looking at samples in a microscope. Conveniently, the body provides its own biological ruler in the form of red blood cells. Red blood cells have a width of about 7 microns and because most samples will have blood vessels, red blood cells are usually available to estimate the size of cells and other structures in histological images.

Epithelia
Introduction
Epithelium is one of the four primary tissues in the human body. Epithelia are composed of sheets of cells that cover most body surfaces and form the cavities and tubes that course throughout our bodies. Epithelia are found along the outer surface of the body (skin), in tracts traversing the body (gastrointestinal tract), in dead-end tracts that have openings at the body surface (respiratory, urinary, and genital tracts), and ducts that open into these tracts (exocrine glands). Because of their wide distribution, the functions of epithelia are numerous, and a single epithelium may have several functions. The most important functions include physical protection and selective transport (diffusion, absorption, secretion) of material.
Basic Structure of Epithelia
Epithelia comprise a continuous sheet of cells that can have one or multiple layers. Cells in an epithelium are linked by several different types of cell junctions that provide physical strength and means of communication. These are discussed in more detail in the material on epithelial structure and transport.
Epithelial usually separate two different compartments. For example, the outside world from our bodies (skin, gastrointestinal tract, respiratory tract) or a fluid from blood (urinary tract). Consequently, epithelia have two surfaces: one that faces the outside world or lumen of a tube and another that faces the underlying tissue. Epithelial polarity is critical for its function. The surface of epithelia that faces the outside world or lumen of a tube we call apical and the side that faces the underlying tissue we call basal.
The basal side of all epithelia attach to a form of extracellular matrix called the basement membrane. The basement membrane provides structural support and separates epithelia from the underlying tissue. Because epithelia lack vasculature (no blood vessels), all nutrients must diffuse from the underlying tissue and across the basement membrane to reach epithelia

Histological Classification of Epithelia
Epithelia are classified based on three criteria:
- Number of Cell Layers: Epithelia with a single layer of cells are called simple epithelia. All of the cells in a simple epithelium contact the basement membrane. Epithelia with more than one layer of cells are called stratified epithelia. In stratified epithelia only the basal layer of cells contacts the basement membrane. A third classification is pseudo-stratified in which the epithelium appears to have multiple layers of cells, but all of the cells contact the basement membrane and therefore, do not form a true stratified epithelia.
- Shape of Cells: Epithelia are also classified based on the appearance of the cells in the outermost layer. If the outermost layer of cells is flattened, the epithelia is called squamous. If the cells in the outer layer are about as tall as they are wide, then the epithelia is called cuboidal. When the cells are taller than they are wide, the epithelia is called columnar. Stratified epithelia are named after the shape of their most superficial layer of cells.
- Surface Specializations: Some epithelial cells contain on their apical surface structures that allow them to perform certain functions.
- Microvilli are small processes that project from the apical surface of many epithelial cells, greatly increasing the surface area. They are notably found on epithelial cells along the intestine and in the proximal convoluted tubule of the kidney.
- Stereo cilia are unusually long, non-motile microvilli that are found on the surface epithelium in the epididymis and ductus deferens.
- Cilia are actively motile processes found on the epithelial lining of the respiratory tract and the female reproductive tract.

Simple Squamous
Simple squamous epithelium is characterized by a continuous surface of irregularly shaped, flat cells supported by an underlying basement membrane. Note the appearance of simple squamous epithelium in section: the flattened nuclei and very thin cytoplasm between the apical and basal surfaces. The thinness of simple squamous epithelium make it ideal for diffusion of gases.

Simple Cuboidal
Simple cuboidal epithelia occur widely in the body in many glands and glandular ducts, such as the salivary ducts, pancreatic duct, bile duct, and kidney tubules. They are often involved in active uptake or secretion of ions, water and proteins. These epithelia are composed of cells that are roughly square with their width and height approximately equal. Their larger size Note that in three dimensions the cells would form a cube. Note that the apical side of the cell faces the lumen.

Simple Columnar
Simple columnar epithelia are similar to cuboidal epithelia but are taller than they are wide. The nuclei are elongated and are often located to one side of the cell. Simple columnar cells are prominent in epithelia where there is active absorption of material (e.g. small intestine, gall bladder). Their larger size allows more room for organelles involved in secretion or absorption of material and generation of ATP. Note the apical surface faces the lumen of the tube.

Simple Columnar with Microvilli
Many cells in a simple columnar epithelium contain microvilli on their apical surface. Microvilli are outward folds of the cell membrane that are supported by parallel arrays of actin filaments. Cutting microvilli in cross-section and examining the cells by electron microscopy reveals the bundles of actin filaments in each microvilli. Microvilli increase the surface area of the apical membrane and make cells more efficient in taking up or releasing material across their apical membrane.

Stratified Squamous
Stratified squamous epithelia have two or more layers of cells, with a squamous layer on the apical surface and basal layers that are usually cuboidal or columnar. The cells in the basal slowly migrate upward toward the surface of the epithelium, and along the way, their nuclei condense and the cells flatten. Eventually, the most superficial layer of cells is sloughed off and replaced by the layer of cells beneath. This type of epithelium can withstand abrasion because the loss of cells from the surface does not compromise the underlying tissue.

Stratified Squamous with Keratin
In this image note the layers of material on the apical surface of the epithelium. This is keratin which is prominent on the the apical surface of the epithelia of skin. Keratin protects against desiccation and abrasion and is composed of a combination of intermediate filaments and lipid. Keratin will be discussed in more detail in the lab on skin.

Cilia
Cilia are long extensions of the apical cell membrane. Cilia are composed of microtubules and the motor protein dynein. Dynein slides the microtubules back and forth within the cilia, creating a wave-like motion of the cilia. The synchronized beating of cilia is capable of moving large objects along the surface of an epithelium. Not that cilia are much longer than microvilli.

Pseudo-stratified
Cilia are often found on epithelia classified as pseudo-stratified. In pseudo-stratified epithelia, the cells become so crowded that the nuclei become displaced and form several rows in the epithelium, making the epithelium appear to be stratified. However, all of the cells in a pseudo-stratified epithelium contact the basement membrane. In a true stratified epithelium only the basal layer of cells contacts the basement membrane. Pseudo-stratified epithelia are most commonly found along the respiratory airways.

Stereocilia
Although stereocilia share part of their name with cilia, they are very different structures. Stereocilia contain actin filaments instead of microtubules and are not motile. Stereocilia facilitate absorption of material and are prominent in the male reproductive tract.

Transitional
Transitional epithelia is a specialized form that is found in the bladder. The hallmark of transitional epithelia is that the shape of the cells and number of layers of the cells depends upon the state of the bladder. When the bladder is filled with urine and becomes distended, the epithelia stretches to accommodate the increase in volume of the bladder and the cells become flatter with fewer layers. In an empty bladder, the decrease in volume of the bladder allows the epithelia to relax and cells become rounder and pile up into more layers.

Basement Membrane
Underlying most epithelia is a basement membrane (basal lamina) which is type of extracellular matrix. The basement membrane provides structural and metabolic support to epithelia. It is usually 30 - 60 nm thick and made up of a network of collagenous and non-collagenous glycoproteins and proteoglycans. Integrins on the basal surface of epithelial cells interact with proteins of the basement membrane. This layer can best be seen in samples stained with the periodic acid Schiff (PAS) procedure which labels carbohydrates which are abundant in the basement membrane.
